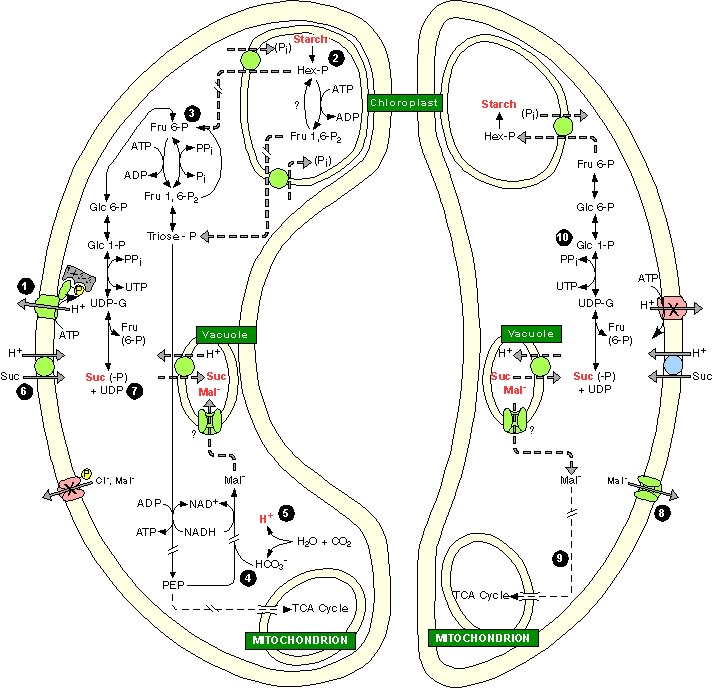
GC metabolism during stomatal opening (left) and stomatal closing (right). Much is unknown about GC carbon metabolism and some processes are inferred. In essence, stomatal opening is initiated by H+ extrusion (1). Because the cytosolic [H+] is low (10-7–10-8 M) and weakly buffered (~10 mM·pH-1), H+ extrusion must be supported by new cytosolic H+ sources, such as H+-Cl- symport (3 in Fig. 1) or release of H+ during A- accumulation. In the latter, starch (2) is degraded to hex-P (3), which is exported to the cytosol and, through glycolysis, provides PEP. Glycolytic oxidation of the aldehyde to the acid releases one H+ to the cytosol. PEP is carboxylated (4) with HCO3-, the synthesis of which releases a second H+ (5). Separately, PM hyperpolarization increases the driving force for suc uptake (6), but control mechanisms are unknown. Some suc possibly is formed from starch degradation (7). GC symplastic [suc] increase accounts for a variable portion of the osmolyte requirement to open stomata, which is in opposition to the GC apoplastic [suc] increase that results from transpiration. Stomatal closing is correlated with mal- efflux (8) or oxidative decarboxylation (9); evidence is against gluconeogenesis. The fate of suc during stomatal closure is unknown, but it is possibly converted to starch (10). Compartments are not to scale.